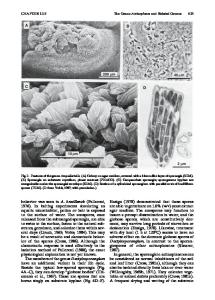
Oceanospirillum and Related Genera
- PDF / 659,648 Bytes
- 29 Pages / 539 x 751 pts Page_size
- 103 Downloads / 313 Views
CHAPTER 3.3.33 mu l l i r i psonaecO
dna
de t a l eR
areneG
Oceanospirillum and Related Genera JOSÉ M. GONZÁLEZ AND WILLIAM B. WHITMAN
Introduction Gram-negative, marine and/or halophilic aerobes and facultative anaerobes belong to several phylogenetic groups. Within the γ-subdivision of the Proteobacteria, a number of well known genera belonging to this group include Alteromonas, Vibrio, Shewanella and Photobacterium. This chapter covers many of the other, lesser known marine genera of the class γ-Proteobacteria, including Alcanivorax, Fundibacter, Hahella, Marinobacter, Marinobacterium, Marinomonas, Marinospirillum, Microbulbifer, Neptunomonas, Oceanobacter, Oceanospirillum and Pseudospirillum, as well as some related organisms that have not been properly classified, such as Pseudomonas halophila and strain 2-40. Historically, many of the species in this group have been classified with Oceanospirillum. Phylogenetic analyses of the 16S rRNA of these taxa suggest that, while these genera are all related, they are also fairly diverse and likely to represent a number of relatively deep phylogenetic groups (Fig. 1). Affiliated with these lesser known genera are the members of the family Halomonadaceae and the Pseudomonas spp. sensu stricto, both of which are discussed in other chapters. In addition, the genus Balneatrix is also affilitated with this group. This genus was isolated during a small outbreak of pneumonia and meningitis in humans in a spa therapy center in 1987 (Dauga et al., 1993) and grows in medium without NaCl. Although Balneatrix and Pseudomonas spp. are not primarily of marine origin, the other members of this group have only been found in marine or halophilic environments, suggesting that there is an important evolutionary distinction between freshwater and marine bacteria. Although not a closely related group, the oceanospirilla have a monophyletic nature which is supported by several types of analyses. Except as noted below, sequence analyses of the 16S rRNA by both neighbor-joining and maximum likelihood methods support the grouping of these genera (Fig. 1). In the case of the maximum likelihood method, all branches are significantly positive (P < 0.01). Bootstrap analyses also indi-
cate that this clade is robust, and it appears in 95% of the replicates. Similarly, the clade containing Shewanella, Vibrio, Alteromonas and the Enterobacteriaceae is clearly resolved. However, this topology is altered greatly when sequences from certain symbionts of marine animals are considered. For example, addition of the sequence of a cellulolytic nitrogen-fixing organism isolated from shipworms (Distel et al., 1991) or the species “Candidatus Endobugula sertula,” a symbiont of the larva of the bryozoan Bugula neritina (Haygood and Davidson, 1997), produces deep branches within the trees and causes the sequences of the Enterobacteriaceae and related families to move inside the oceanospirilla. Because these alternative topologies are only recovered in the presence of certain sequences, they appeared to be artifacts.
Data Loading...